-
Email info@jnans.org
-
Address 848 N. Rainbow Blvd. #5486 Las Vegas, NV 89107, USA
1School of Life Sciences, Westlake University, Hangzhou, China.
2College of Biomedical Engineering and Instrument Science, Zhejiang University, Hangzhou, China.
3Molemuse Biotech Studio, Xihu District, Hangzhou, China.
*Corresponding author: Fangfei Zhang
Molemuse Biotech Studio, Xihu District, Hangzhou, China.
Email ID: zhangfangfei@westlake.edu.cn
Received: Jun 24, 2025
Accepted: Jul 23, 2025
Published Online: Jul 30, 2025
Journal: Journal of Neurology and Neurological Sciences
Copyright: Zhang F et al. © All rights are reserved
Citation: He S, Hu Y, Zhang F. Integrative analysis of microRNA as blood-based biomarkers in amyotrophic lateral sclerosis. J Neurol Neuro Sci. 2025; 1(2): 1009
Amyotrophic Lateral Sclerosis (ALS) is a fatal neurodegenerative disease with limited diagnostic and prognostic tools. The identification of minimally invasive biomarkers is critical for early diagnosis, monitoring disease progression, and evaluating therapeutic responses. MicroRNAs (miRNAs), small non-coding RNAs that regulate gene expression, have emerged as promising blood-based biomarkers due to their stability and diseasespecific expression patterns. Numerous studies have reported dysregulated circulating miRNAs in ALS patients, including miR-206, miR-133b, and miR-338-3p, some of which correlate with clinical parameters such as disease severity and survival. However, methodological variability across studies has led to inconsistent findings, hindering clinical translation. In this study, we conducted a systematic review and integrative analysis of blood-derived miRNA biomarkers in ALS. Through manual curation of published literature, we identified consistently dysregulated miRNAs and assessed their associations with clinical outcomes. Using bioinformatics tools, including STREME, TargetScanHuman 8.0, and miRPath v4.0, we performed motif enrichment, target prediction, and pathway analysis to explore the functional relevance of these miRNAs in ALS pathogenesis. Our findings highlight potential miRNA candidates with diagnostic and prognostic value and provide insights into the molecular mechanisms underlying ALS. This integrative approach offers a foundation for future validation studies and the development of standardized protocols for miRNA-based biomarker discovery in ALS.
Amyotrophic Lateral Sclerosis (ALS) is a progressive neurodegenerative disorder characterized by the loss of motor neurons, leading to muscle weakness, paralysis, and eventual respiratory failure. Despite its devastating impact, the pathogenesis of ALS remains incompletely understood, and no disease-modifying therapies currently exist. A critical barrier to progress is the absence of reliable, minimally invasive biomarkers for early diagnosis, disease progression tracking, and therapeutic response monitoring [1]. While Cerebrospinal Fluid (CSF) and tissuebased biomarkers have shown promise, their invasive collection procedures and limited dynamic monitoring capacity hinder clinical utility. Blood-based biomarkers, by contrast, offer accessibility, repeatability, and potential for integration into routine clinical care [2].
Among emerging candidates, microRNAs (miRNAs)—small, non-coding RNAs that regulate gene expression post-transcriptionally—have garnered significant attention [3]. miRNAs are stable in blood, encapsulated in exosomes or bound to proteins, and can reflect cellular pathology even in remote tissues. In ALS, dysregulation of specific miRNAs has been implicated in motor neuron degeneration, neuroinflammation, and oxidative stress, suggesting their potential as disease-specific biomarkers [4]. Numerous studies have reported altered circulating miRNA profiles in ALS patients compared to healthy controls or neurodegenerative disease controls, with some miRNAs (e.g., miR206 [5-7], miR-133b [6,7], miR-338-3p [8]) consistently associated with disease activity or survival.
However, a critical gap persists in the field: despite a surge of individual biomarker studies, there has been no comprehensive, integrative analysis of these findings.
Methodological heterogeneity—including variations in sample collection, miRNA isolation techniques, normalization strategies, and statistical approaches—has led to inconsistent results across studies. For instance, some reports highlight miR-206 as a marker of muscle atrophy, while others find no correlation with clinical parameters. Additionally, small sample sizes, lack of longitudinal data, and inconsistent control groups (e.g., healthy controls vs. disease controls) further complicate comparisons. This fragmentation has hindered the translation of miRNA biomarkers into clinical practice and obscured their true diagnostic and prognostic potential.
This work aims to systematically summarize and critically evaluate the current literature on ALS blood-based miRNA biomarkers. By integrating findings from diverse studies, we seek to identify consistently dysregulated miRNAs, assess their correlation with clinical outcomes (e.g., ALS Functional Rating Scale scores, survival), and explore methodological factors contributing to variability. Through meta-analysis and quality assessment, we aim to refine candidate biomarkers and propose standardized protocols for future validation. Ultimately, this integrative approach could pave the way for robust, blood-based miRNA biomarkers that enhance ALS diagnosis, stratify patients for clinical trials, and monitor disease progression—bridging the critical gap between basic research and clinical translation.
Literature search
To identify studies on blood-based microRNA (miRNA) biomarkers in Amyotrophic Lateral Sclerosis (ALS), a systematic search of PubMed was conducted using the query (biomarker OR biomarkers OR biomarker) AND (amyotrophic lateral sclerosis) AND (“blood” OR “plasma” OR “serum”), yielding 979 articles. These were manually curated by two independent reviewers to select clinically relevant studies investigating miRNA expression in ALS blood, plasma, or serum. Articles were excluded if they focused on non-human models, lacked miRNAspecific data, or did not report associations between miRNA levels and ALS clinical parameters (e.g., diagnosis, progression, survival). This structured approach ensured the selection of high-quality evidence for integrative analysis.
Literature annotation
Manual checks were performed for each publication identified in PubMed. Kimi Explorer (version 1.1.2), a Microsoft Edge extension, was employed with the prompt: “Summarize the changed miRNA in this format: miRxxx (1, indicating upregulated/-1, indicating downregulated). Collect the toolkit names for miR extraction and measurements and indicate sample source and cohort arrangements. Finally, check whether the data is open or not.”
The results were structured to reflect tissue-specific miRNA regulation, methods for miRNA detection (e.g., qRT-PCR, microarrays), specific controls (e.g., endogenous housekeeping genes, spike-in RNAs), and data availability (publicly accessible via repository links, if applicable) were documented. Patient cohort details, including demographic origin (e.g., geographic region, clinical setting) and group stratification (e.g., case-control, age-matched subgroups), were summarized. The compiled data were organized into an Excel spreadsheet, and WPS AI’s “translate to English” feature ensured consistent formatting and language uniformity.
RNA sequence motif enrichment
miRNA sequence motif enrichment analysis using STREME: To identify enriched sequence motifs in the dysregulated miRNAs, the top differentially expressed miRNAs were analyzed using the STREME (Short Time-series Motif Enrichment) web tool https://meme-suite.org/meme/tools/streme, which is part of the MEME Suite. The input sequences consisted of the mature miRNA sequences obtained from miRBase. STREME performs unbiased motif discovery by scanning the input sequences for significantly overrepresented short motifs, and compares them against a background set of sequences — typically all expressed miRNAs or randomly selected miRNAs from the same species.
miRNA target prediction using TargetScanHuman 8.0: To predict potential target genes of the dysregulated miRNAs, TargetScanHuman 8.0 was employed. This computational tool utilizes evolutionary conservation and seed-region matching to identify putative binding sites within the 3’ Untranslated Regions (UTRs) of target mRNAs. The analysis was performed using the default parameters, focusing on conserved targeting across species. Predicted targets were filtered based on context++ scores, which provide a quantitative measure of the likelihood that a given miRNA–mRNA interaction is functional.
miRNA pathway analysis using miRPath v4.0: An miRNAcentric pathway analysis was performed using miRPath v4.0, a computational tool designed to explore the functional roles of miRNAs in biological pathways. The analysis was conducted using the top dysregulated miRNAs identified from the datasets. Experimentally validated miRNA-target interactions were obtained from TarBase v8.0, while miRNA annotations were retrieved from miRBase v22.1. In this approach, miRPath v4.0 was used to predict enriched pathways based on the experimentally supported target genes of the selected miRNAs. Pathway enrichment was assessed using the DIANA algorithm and the Kyoto Encyclopedia of Genes and Genomes (KEGG) database. Statistical significance was determined using a False Discovery Rate (FDR)-corrected p-value threshold.
Overview of the dysregulated miRNA
To gain insights into the expression patterns and potential functional relevance of dysregulated miRNAs, a systematic review of the retrieved PubMed literature was conducted, focusing on clinical relevance and study type. This process resulted in a final collection of 28 studies [9-36] (Figure 1), which were manually curated to extract information regarding the direction of miRNA regulation—specifically, whether each miRNA was reported as upregulated or downregulated. Key experimental details, including sample type, patient cohort size, and miRNA isolation methods, were also recorded for each study (Table 1).
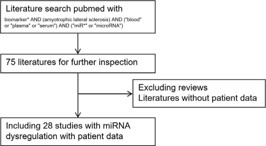
For each study, the total number of dysregulated miRNAs was summarized and categorized based on the sample type (Figure 2A). Furthermore, for each individual miRNA, the frequency of its differential expression—either up or down—was compiled across all included studies (Figure 2B). This synthesis aims to highlight miRNAs that are recurrently deregulated under similar conditions, offering a foundation for further exploration of their potential roles in disease mechanisms or as candidate biomarkers.
Additionally, the miRNA extraction methods employed in each study were recorded and categorized, allowing for an overview of the most commonly used techniques in miRNA isolation (Figure 2C). We also summarized the distribution of patient cohort sizes across the included studies, ranging from small-scale investigations to larger clinical cohorts (Figure 2D). This information provides insight into the variability in sample processing and potential influences on miRNA detection consistency across different experimental settings.
Characterization of the dysregulated miRNA
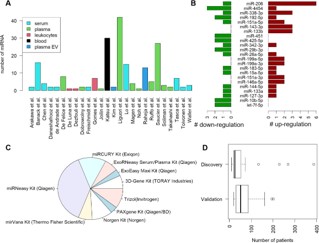
The dysregulated miRNAs were categorized into upregulated and downregulated groups for downstream functional analysis. The upregulated group included the following miRNAs: miR206, miR-4454, miR-338-3p, miR-192-5p, miR-151a-5p, miR-143-3p, miR-133b, miR-342-3p, miR-26a-5p, miR-199a-5p, miR199a-3p, miR-183-5p, miR-15a-5p, miR-151a-3p, miR-146a-5p, miR-144-5p, miR-133a, and miR-127-3p. In contrast, the downregulated group comprised miR-206, miR-4454, miR-338-3p, miR-192-5p, miR-151a-5p, miR-451, miR-425-5p, miR-342-3p, miR-29b-3p, miR-26a-5p, miR-183-5p, miR-15a-5p, miR-1445p, miR-133a, miR-127-3p, miR-10b-5p, and let-7f-5p. These two sets of miRNAs were further subjected to sequence motif enrichment analysis using their corresponding FASTA sequences to identify potential shared regulatory elements or functional patterns (Figure 3). In the upregulated group, a conserved motif CCAGG was identified as significantly enriched (Figure 3A). This motif may represent a key regulatory element that contributes to the coordinated expression or functional activity of these miRNAs. It could serve as a binding site for transcription factors or RNA-binding proteins that influence miRNA biogenesis, stability, or target specificity. The presence of this motif suggests a possible shared regulatory mechanism among the upregulated miRNAs, offering new insights into their expression control and biological roles in the studied condition.
Enrichment of the pathway regulated by dysregulated miRNA
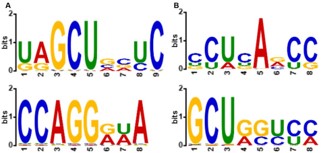
To elucidate the biological pathways potentially regulated by dysregulated miRNAs in ALS, we selected miRNAs that were reported as either upregulated or downregulated in at least two independent studies. These miRNAs were subjected to target prediction using TarBase 8.0, followed by pathway enrichment analysis via KEGG using miRPath v4.0.
As illustrated, the most significantly enriched pathways include “Proteoglycans in cancer,” “Ubiquitin mediated proteolysis,” and the “p53 signaling pathway,” Each Demonstrating high-log10(FDR) values, indicative of strong statistical significance. Additional pathways of interest encompass “Protein processing in endoplasmic reticulum,” “Cell cycle,” and several signaling pathways such as “Hippo,” “FoxO,” and “AMPK.” Notably, pathways related to neurodegeneration, including “Amyotrophic lateral sclerosis,” “Alzheimer disease,” and “Pathways of neurodegeneration multiple diseases,” were also significantly enriched, underscoring the relevance of these miRNAs to ALS pathogenesis.
These findings suggest that dysregulated miRNAs in ALS may exert their effects through a diverse array of biological processes, including cell cycle regulation, protein homeostasis, and canonical cancer and neurodegeneration-related pathways. The enrichment of both general and disease-specific pathways highlights the complex molecular landscape influenced by miRNA dysregulation in ALS.
| Publication | PMID | Dysregulated MicroRNAs | Sample Origin | Discovery Cohort ALS | Discovery Cohort Control | Validation Cohort ALS | Validation Cohort Control | RNA extraction Method |
|---|---|---|---|---|---|---|---|---|
| Freischmidt et al. | 26142125 | miR-1234-3p (-1), miR-1825 (-1) | serum | 18 | 16 | 20 | 13 | |
| Tasca et al. | 26588026 | miR-206 (1), miR-133a (1), miR-133b (1), miR-146a (-1), miR-149* (-1), miR-155 (-1), miR-27a (-1) | serum | 14 | 8 | miRNeasy Kit (Qiagen) | ||
| Waller et al. | 28454844 | miR-206 (1), miR-143-3p (1), miR-374b-5p (-1) | serum | 27 | 25 | 23 | 22 | Norgen Kit (Norgen) |
| Raheja et al. | 29466830 | miR-1 (1), miR-133a-3p (1), miR-133b (1), miR-144-5p (1), miR-192-3p (1), miR-192-5p (1), miR-19a-3p (1); miR-320c (-1), miR-320a (-1), let-7d-3p (-1), miR-425-5p (-1),miR-320b (-1), miR-139-5p (-1) | serum | 23 | 30 | miRCURY Kit (Exiqon) | ||
| Kim et al. | 37016037 | miR-23c (1), miR-192-5p (-1) | serum | 12 | 11 | 18 | 15 | ExoEasy Maxi Kit (Qiagen) ExoRNeasy Serum/Plasma Kit (Qiagen) |
| Banack et al. | 32574550 | miR-146a-5p (1), miR-199a-3p (1), miR-4454 (-1), miR-10b-5p (-1), miR-29b-3p (-1), miR-151a-3p (1), miR-151a-5p (1), miR-199a-5p (1) | plasma | 10 | 10 | 10 | 10 | ExoRNeasy Serum/Plasma Kit (Qiagen) |
| De Felice et al. | 22903028 | miR-451 (-1), miR-1275 (-1), miR-328-5P (-1), miR-638 (-1), miR-149 (-1), miR-665 (-1), miR-338-3p (1) | leukocytes | 8 | 12 | 14 | 14 | Trizol(Invitrogen) |
| De Felice et al. | 25130371 | miR-338-3p (1) | leukocytes | 72 | 62 | 10 | 10 | Trizol(Invitrogen) |
| de Andrade et al. | 27538595 | miR-424 (1), miR-206 (1) | plasma | 5 | 5 | 39 | 39 | miRNeasy Kit (Qiagen) |
| Toivonen et al. | 24586506 | miR-206 (1), miR-106b (1) | serum | 12 | 12 | Norgen Kit (Norgen) | ||
| Chen et al. | 27582688 | miR-183 (-1), miR-193b (-1), miR-451 (-1), miR-3935 (-1) | leukocytes | 5 | 5 | 83 | 61 | miRNeasy Kit (Qiagen) |
| Takahashi et al. | 26497046 | miR-4649-5p (1), miR-4299 (-1) | plasma | 16 | 10 | 48 | 47 | 3D-Gene Kit (TORAY Industries) miRNeasy Kit (Qiagen) |
| Liguori et al. | 30210287 | let-7a-5p (-1), let-7d-5p (-1), let-7f-5p (-1), let-7g-5p (-1), let-7i-5p (-1), miR-103a-3p (-1), miR-106b-3p (-1), miR-128-3p (-1), miR-130a-3p (-1), miR-130b-3p (-1), miR- 144-5p (-1), miR-148a-3p (-1), miR-148b-3p (-1), miR-15a-5p (-1), miR-15b-5p (-1), miR-151a-5p (-1), miR-151b (-1), miR-16-5p (-1), miR-181a-2-3p (-1), miR-182-5p (-1), miR-183-5p (-1), miR-186-5p (-1), miR-192-5p (-1), miR-22-3p (-1), miR-221-3p (-1), miR-223-3p (-1), miR-23a-3p (-1), miR-25-3p (-1), miR-26a-5p (-1), miR-26b-5p (-1), miR-27b-3p (-1), miR-28-3p (-1), miR-30b-5p (-1), miR-30c-5p (-1), miR-342-3p (-1), miR-409-3p (-1), miR-425-5p (-1), miR-451a (-1), miR-532-5p (-1), miR-550a-3p (-1), miR-584-5p (-1), miR-93-5p (-1) |
blood | 6 | 5 | 50 | 15 | PAXgene Kit (Qiagen/BD) |
| Katsu et al. | 31173847 | miR-4736 (1), miR-4700-5p (1), miR-1207-5p (1), miR-4739 (1), miR-4505 (1), miR-24-3p (1), miR-149-3p (1), miR-4484 (1), miR-4688 (1), miR-4298 (1), miR-939-5p (1), miR-371a-5p (1), miR-3619-3p (1), miR-1268a (-1), miR-2861 (-1), miR-4508 (-1), miR-4507 (-1), miR-3176 (-1), miR-4745-5p (-1), miR-3911 (-1), miR-3605-5p (-1), miR- 150-3p (-1), miR-3940-3p (-1), miR-4646-5p (-1), miR-4687-5p (-1), miR-4788 (-1), miR-4674 (-1), miR-1913 (-1), miR-634 (-1), miR-3177-3p (-1) |
plasma EV | 5 | 5 | 3D-Gene Kit (TORAY Industries) | ||
| Magen et al. | 34711961 | miR-423 (1), miR-484 (1), miR-92a (1), miR-92b (1) | plasma | 126 | 103 | 122 | 73 | miRNeasy Kit (Qiagen) |
| Lo et al. | 34776863 | miR-1254 (-1), miR-587 (-1), miR-766-3p (-1), miR-298 (-1), miR-877-5p (-1), miR-26a-5p (1), miR-1262 (1), miR-551b-3p (1), miR-1268b (1), miR-450a-2-3p (1), miR-127-3p (1), miR-7975 (1), miR-4454 (1), miR-520f-3p (1), miR-342-3p (1), miR-1255a |
serum | 15 | 16 | Norgen Kit (Norgen) | ||
| Gomes et al. | 37531027 | miR-7-2-3p (1), miR-26a-1-3p (1), miR-224-5p (1), miR-206 (1), miR-361-5p (-1), miR-3159 (1), miR-630 (-1) | plasma | 15 | 16 | 35 | 16 | miRNeasy Kit (Qiagen) |
| Noh et al. | 39915090 | miR-214 (1) | plasma | 15 | 5 | 132 | 30 | |
| Banack et al. | 39280119 | miR-10b-5p (-1), miR-4454 (-1), miR-199a-3p (1) , miR-151a-3p (1), miR-151a-5p (1), miR-199a-5p (1), miR-146a-5p (1), miR-29b-3p (-1) | plasma EV | 119 | 150 | miRNeasy Kit (Qiagen) | ||
| Dezfouli et al. | 40097890 | miR-223 (-1) | plasma | 32 | 32 | miRNeasy Kit (Qiagen) | ||
| Ruffo et al. | 37189452 | miR-143-3p (1), miR-574-5p (1), miR-133b (1), let-7b-5p (-1), miR-146a-3p (-1) | plasma | 7 | 13 | 7 | 13 | mirVana Kit (Thermo Fisher Scientific) |
| Daneshafrooz et al. | 35082326 | let-7f-5p (-1), miR-338-3p (1) | plasma | 206 | 181 | 30 | 30 | Trizol(Invitrogen) |
| Soliman et al. | 34454204 | miR-206 (1), miR-143-3p (1), miR-142-3p (1) | plasma | 30 | 20 | miRNeasy Kit (Qiagen) | ||
| Dobrowolny et al. | 33431881 | miR-206 (-1), miR-133a (-1), miR-151a-5p (-1) | serum | 13 | 6 | 19 | 4 | miRNeasy Kit (Qiagen) and miRCURY Kit (Exiqon) |
| Arakawa et al. | 32739158 | miR-92a-3p (1), miR-486-5p (1) | serum | 5 | 16 | 14 | 25 | miRCURY Kit (Exiqon) |
| Joilin et al. | 32613197 | miR-21-5p (-1) | serum | 48 | 24 | 156 | 46 | miRNeasy kit (Qiagen) |
| De Luna et al. | 32152380 | miR-335-5p (-1) | serum | 7 | 6 | 53 | 23 | miRCURY Kit (Exiqon) |
| Saucier et al. | 30552897 | miR-532-3p (1), miR-144-3p (1), miR-15a-5p (1), miR-363-3p (1), miR-183-5p (1), miR-4454 (-1), miR-9-1-5p (-1), miR-9-3-5p (-1), miR-338-3p (-1), miR-9-2-5p (-1), miR-100-5p (-1), miR-7977 (-1), miR-1246 (-1), miR-664a-5p (-1), miR-7641-1 (-1), miR-1290 (-1), miR-4286 (-1), miR-181b-1-5p (-1), miR-1260b (-1), miR-181b-2-5p (-1), miR- 127-3p (-1), let-7c-5p (-1), miR-181a-2-5p (-1), miR-181a-1-5p (-1), miR-199a-2-3p (-1), miR-199b-3p (-1), miR-199a-1-3p (-1) |
plasma EV | 14 | 12 | mirVana Kit (Thermo Fisher Scientific) |
MicroRNAs (miRNAs) have emerged as important regulators in ALS, influencing cell fate, promoting tumor growth, and holding therapeutic potential. However, consistent patterns of miRNA dysregulation in ALS remain elusive. This inconsistency largely stems from technical challenges, including the lack of comprehensive whole-miRNA profiling, which hampers effective internal normalization and prevents integration of datasets across studies. Additionally, the short length of miRNAs complicates primer design for qPCR, limiting specificity and sensitivity. To overcome these limitations, In Situ Hybridization (ISH) with locked nucleic acid probes offers a valuable approach for native level validation of dysregulated miRNAs in patient samples.
Another major obstacle is the lack of standardization in patient cohort selection and sample sources, which restricts the generalizability of miRNA biomarker findings. Although community efforts like the ALL ALS and Project MinE consortia have advanced genetic and proteomic profiling in ALS, miRNA research remains underrepresented. Integrating miRNA profiling into these large-scale initiatives, including iPSC-based models, could provide deeper mechanistic insights and improve biomarker discovery.
To advance the field, future research must focus on standardized protocols for sample collection and miRNA analysis, improved detection methods, and collaborative data sharing platforms. Raising awareness of miRNA’s critical role in ALS will encourage the research community to address these challenges, ultimately facilitating the development of reliable miRNA biomarkers and therapeutic targets for this devastating disease.
In conclusion, our systematic meta-analysis demonstrates that circulating microRNAs represent promising blood-based biomarkers for ALS diagnosis, despite the inherent heterogeneity that characterizes both the disease and biomarker expression patterns. The identification of consistent dysregulation patterns for key microRNAs across multiple independent studies provides strong evidence for their clinical utility [37]. While methodological standardization and large-scale validation studies remain essential for clinical translation, the mechanistic relevance and diagnostic potential of these biomarkers position them as valuable tools for improving ALS diagnosis and patient care [38].
The observed heterogeneity in microRNA expression patterns, rather than representing a limitation, may actually reflect the biological complexity of ALS and provide opportunities for patient stratification and personalized medicine approaches [39]. Future research incorporating comprehensive microRNA panels, standardized methodologies, and integration with clinical parameters will be crucial for realizing the full potential of these biomarkers in ALS diagnosis and management [40,41].
This integrative analysis contributes significantly to the growing body of evidence supporting microRNA biomarkers in neurodegenerative diseases and provides a foundation for their continued development as clinically relevant diagnostic tools for ALS [42]. The convergence of molecular insights, diagnostic utility, and mechanistic understanding positions circulating microRNAs as a promising frontier in ALS biomarker research and clinical application [43].
